Hui Tang ![]() ,
Jun Wang,
Lei Zhao,
Xue-mei Zhao
,
Jun Wang,
Lei Zhao,
Xue-mei Zhao
For correspondence:- Hui Tang Email: tanghui494@126.com Tel:+8653187938925
Received: 6 November 2016 Accepted: 20 February 2017 Published: 31 March 2017
Citation: Tang H, Wang J, Zhao L, Zhao X. Rhodiola rosea L extract shows protective activity against Alzheimer's disease in 3xTg-AD mice. Trop J Pharm Res 2017; 16(3):509-514 doi: 10.4314/tjpr.v16i3.3
© 2017 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the protective effect of Rhodiola rosea L. extract (RRLE) against Alzheimer's disease in 3xTg-AD mice.
Methods: The cognitive function of 3xTg-AD mice was assessed using Morris water maze test. The levels of amyloid beta deposits and NeuN in the hippocampus were evaluated by immunohistochemistry. Brain neurotrophic-derived factor (BDNF) and tyrosine kinase B (TrkB) ex
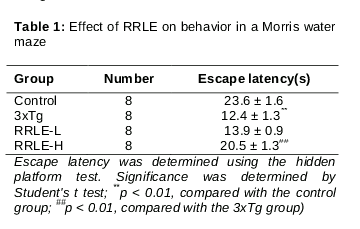
Results: RRLE treatment significantly ameliorated learning and memory deficits in AD mice, as shown by increased time spent in the target zone during probe tests. The escape latency in animals treated with 400 mg/kg RRLE (20.5 ± 1.3 s) was significantly increased compared to the untreated mice (12.4 ± 1.3 s, p < 0.01). In addition, RRLE significantly decreased Aβ deposits, increased NeuN-positive cells, and upregulated the ex
Conclusion: The findings suggest that RRLE treatment may be a useful strategy for treating memory impairment induced by several neurodegenerative diseases
Introduction
Alzheimer’s disease (AD) is the most common form of dementia in the elderly and accounts for between 50 and 75 % of all cases. By 2030 it is estimated that more than 65 million people will be living with dementia, with projections almost doubling every 20 years. AD is a progressive neurodegenerative disorder characterized, at least in part, by abnormal accumulation of β-amyloid peptide (Aβ) in the brain [1]. The accumulated Aβ is believed to play an important role in the pathogenesis of AD [2]. Thus, Aβ continues to be an important target for prevention and treatment of AD [3].
Alzheimer's disease is the most common form of dementia relating to memory and cognitive decline. AD is a progressive neurodegenerative disease in which dementia symptoms gradually worsen over a number of years [4]. Classical biochemical hallmarks of AD include the accumulation of amyloid beta (Aβ) peptide oligomers and soluble hyperphosphorylated tau [5]. Brain-derived neurotrophic factor (BDNF) is a small dimeric protein, and BDNF acts through high affinity binding with its receptor, tyrosine kinase B (TrkB). BDNF modulates neuronal growth and survival, and BDNF is implicated in learning and memory processes; therefore, dysfunction in BDNF is accompanied by cognitive deficits. In particular, BDNF is involved in the AD-related decline of neurogenesis, and levels of nerve growth factor are also diminished with AD [6].
Medical treatment for AD patients is placing an increasing burden on physicians and families every year. Clinically, there are a variety of drugs available for AD, such as cholinesterase inhibitors, glutamate receptor antagonists, and free radical scavengers. However, these drugs do not closely target the pathogenesis of the disease and have numerous side effects [7]. Therefore, it is extremely important to elucidate the mechanism of AD pathophysiology to find a new drug for treatment.
Rhodiola rosea L., has been used in China in the treatment of various disorders, including stress-induced physiological changes, inflammation, hypertension, and cancer. Rhodiola rosea L. extract has been reported to have anti-oxidant [8], immunomodulatory [9], and anti-mutagenic activity [10]. In this study, we investigated to evaluate the neuroprotective effects of RRLE on learning and memory deficits in a triple-transgenic mouse model of Alzheimer's disease (3xTg-AD), which expresses APPSwe, PS1M146V, and tauP301L [11].
Methods
Plant material
The plant material, Rhodiola rosea L., were collected from Pingdingshan City, Henan Province in China in October 2015. Taxonomic identification of the plant was performed by Professor Gang Wang of Shandong University, in China. A voucher specimen (no. PRLE 201605002) was deposited in College of Pharmacy, Shandong University, China for future reference.
The herbal samples Rhodiola rosea L. was dried in an oven. RRLE was obtained by steeping the dried Rhodiola rosea L. in water at 60 oC three times, each for one hour before first drying in an oven and then freeze-drying the RRLE thus obtained. One gram powder was equivalent to about 1.7 g crude samples. Thus, the yield was 58.82 %.
Animals
Alzheimer's disease (3xTg-AD) mice carrying a mutant APP (KM670/671NL), a human mutant PS1 (M146V) knock-in, and tau (P301L) transgenes (B6;129-Psen1tm1Mpm Tg (APPSwe,tauP301L)1Lfa/J) were purchased from the animal research institute, Nanjing University (Nanjing, China). The non-transgenic littermates were used as wild type (WT) controls. All animals were kept in a pathogen-free environment on a 12 h light/dark cycle and had access to food and water ad libitum.
Animal groups
The mice were randomly divided into four groups as follows: (1) saline treated WT group (WT, n = 8); (2) saline-treated 3xTg group (3xTg, n = 8); (3) 200 mg/kg RRLE-treated 3xTg group (3xTgþ + RRLE 200, n = 8) and (4) 400 mg/kg RRLE-treated 3xTg group (3xTgþ + RRLE 400, n = 8). Starting at 3 months of age, mice received PBS and RRLE once a week for 3 months until they were 6 months old. Drugs were dissolved in water, and administered using a 5-ml syringe with a 2-cm long gavage needle through the mouth to the mouth once daily for 3 weeks. The animal experiment was approved by the Animal Care and Use Committee of Shandong University (approval ref no. 20100405) and was carried out in compliance with the Directive 2010/63/EU on the handling of animals used for scientific purposes [12].
Water maze test
A modified version of the water maze procedure described by Morris was used to test each mouse's cognitive function [13]. The water maze was a circular pool 0.9 m in diameter and constructed of fiberglass. Water in the pool was maintained at 22 ± 2 oC and mixed with 1 kg of powdered skim milk to make the water opaque. During testing in the water maze, a platform (6 cm in diameter) was fixed 1 cm below the surface of the water at identical location within the pool. The pool was surrounded by differing extra-maze cues. All mice were subjected to four trials per day at intervals of 15 min for four consecutive days. The proportion of time spent searching for the platform in the training quadrant, i.e., the previous location of the platform, was used as a measure of memory retention.
Western blotting
At the end of the experiment, the mice were decapitated and the brains were rapidly removed and placed on ice. The hippocampus was quickly dissected by a scalpel and stored at -80 oC fridge until use. The hippocampal tissue was homogenized in PRO-PREPTM Protein Extraction Solution (Shanghai Shengong, Shanghai, China). The homogenates were subsequently centrifuged at 12,000 g for 10 min at 4 oC, and the supernatants were collected for protein concentration determination using a protein assay (Bio-Rad, Hercules, CA, USA). Protein samples (30 μg) were separated on a sodium dodecyl sulfate-polyacryl-amide gel and transferred onto a nitrocellulose membrane. The membranes were incubated with 5 % skim milk in Tris-buffered saline containing 0.1 % Tween-20 and then incubated overnight at 4 oC with the following primary antibodies: mouse β- actin antibody (1 : 1000; Santa Cruz Biotechnology, Santa Cruz, CA, USA), rabbit BDNF antibody (1 : 500; Santa Cruz Biotechnology), and rabbit TrkB antibody (1 : 1000; Santa Cruz Biotechnology). Subsequently, membranes were incubated for 1 h with secondary antibodies (1 : 2000; Cell Signaling), and detection was performed using the enhanced chemiluminescence (ECL) detection kit.
Densitometry analysis
Coronal sections of the hippocampus were examined from the rostral anteroposterior (- 2.1 mm) to the anteroposterior (- 4.5 mm) direction, as defined by the bregma of the brain atlas. Images were obtained at 10 × magnification using the IMAGE PRO PLUS System (version 4.0; Media Cybernetics, Silver Spring, MD, USA) on a computer attached to a light microscope (Zeiss Axioskop, Oberkochen, Germany), which interfaced with a chargecoupled device video camera (Kodak Mega Plus model 1.4 I). To determine the density of the Aβ-immunoreactive staining in the hippocampus, a square frame of 500 × 500 μm2 was placed in the dorsal part of the hippocampus. A second square frame of 200 × 200 μm2 was placed in the corpus callosum to measure the background. As previously described, variations in background illumination were controlled by subtracting the average background density of the corpus callosum from the average density of the hippocampus in each section analyzed [14].
Statistical analysis
Data are presented as mean ± standard deviation (SD). Results were analyzed statistically by one-way ANOVA followed by Tukey’s multiple comparison using SPSS 16.0 software for Windows. Differences were considered statistically significant at p < 0.05.
Results
RRLE reverses spatial learning deficits at 6 months of age in mice
To examine spatial memory, the performance of animals in the probe trial was compared to the time animals spent swimming to the platform. All animals were examined by a retention test that involved removing the platform from the pool on the fourth trial day. From , the escape latency in animals treated with 400 mg/kg RRLE was significantly increased as compared to the 3xTg-AD mice.
3xTg-AD mice displayed severely impaired spatial cognition in the water maze test compared with the WT controls, and the administration of RRLE ameliorated these learning and memory deficits.
Effect of RRLE on Aβ pathology in mice
In 3xTg-AD mice, Aβ is present in the hippocampus by 6 months of age. Therefore, to investigate the link between neurogenesis and the development of AD pathology, we assessed the Aβ burden in brains from 3xTg-AD mice that began receiving RRLE treatments 3 month prior to 6 months of age. Aβ deposits in the CA1 region of the hippocampus were significantly increased in 6-month-old 3xTg animals compared to the age-matched WT group (p < 0.01). In the CA3 region of the hippocampus, Aβ deposits were significantly increased in the 3xTg group (p < 0.001) compared to the WT group (p < 0.01).
Compared with the 3xTg group, Aβ deposits in the CA1 region were significantly decreased by RRLE 400 mg treatment (p < 0.05). In addition, compared with the 3xTg group, Aβ deposition in the CA3 was significantly decreased by RRLE 200 mg treatment (p < 0.05). However, there were no significant differences between the 3xTg group and the 3xTgþ RRLE (400 mg/kg) ().
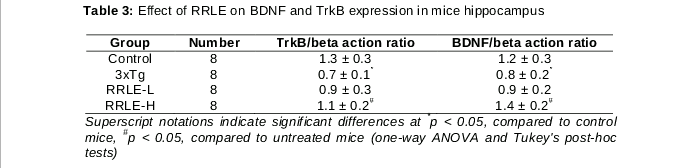
Effect of RRLE on the expressions of BDNF and TrkB and in mice hippocampus
The effect of RRLE treatment on the expression of hippocampal BDNF and TrkB in mice was determined. The results are shown in . The expression of hippocampal BDNF and TrkB of mice treatment with saline was significantly lower compared to that of WT mice (p < 0.05).It was also demonstrated that RRLE treatment significantly increased the expression of hippocampal BDNF and TrkB. These results show that the induction of AD reduced BDNF and TrkB expressions in the hippocampus, whereas, RRLE treatment enhanced BDNF and TrkB expressions in the hippocampus of the 3xTg mice.
Discussion
The present study demonstrated that RRLE increased spatial learning, memory abilities, and the expression of hippocampal BDNF and TrkB in 3xTg-AD mice. In the present study, we chose to use 3xTg-AD mice, a model derived from APPSwe, PS1M146V, and tauP301L transgenes. The 3xTg-AD mice develop a progressive, age-related neuropathological phenotype that includes plaque and tangle pathologies. Both of these hallmark lesions are mainly limited to the hippocampus, amygdala, and cerebral cortex – the brain structures most impacted by AD pathology [15].
Cognitive impairment has been detected in 2-month-old 3xTg-AD mice [16], and Aβ deposits in the hippocampus and cortex have been found in 6-month-old 3xTg-AD mice [17-19]. These findings indicate that the pathological features that imitate AD in 3xTgAD mice remain stable. Classic symptoms of AD include problems with spatial learning and memory deficits. Our study demonstrated that treatment with RRLE resulted in a significant restoration of spatial learning and memory function in AD mice. These results suggest that RRLE treatment may be effective in ameliorating cognitive impairment caused by AD.
Neuron-specific nuclear antigen (NeuN) is a neuronal-specific nuclear protein [20,21]. The expression of NeuN is observed in most neuronal cell types throughout the nervous system, with the exception of some neuronal populations that are NeuN-negative, but does not stain non-neuronal cells [22]. NeuN is a soluble nuclear protein that is localized to the cell nucleus and in the neuronal cytoplasm of postmitotic neurons. Within the hippocampus, NeuN can be used as a marker of postmitotic cells and labels both “normal” postmitotic neurons and newly generated postmitoticneurons. BDNF plays pivotal roles in learning, memory, and neuronal plasticity.
The levels of BDNF, and its main receptor TrkB, have been reported to lessen in AD. It was hypothesized that BDNF and its receptor may be involved in the protective role of RRLE against memory impairment. Results of the present study have demonstrated that RRLE intake significantly increases the expression of BDNF and its main receptor TrkB, in the brain, which is in agreement with our hypothesis. There is evidence to support our results. It has been demonstrated that BDNF and TrkB, are capable of protecting against memory impairment and regulate neurogenesis in the hippocampus of AD. A recent study also supports the role of BDNF signaling through TrkB in the pathophysiology and cognitive and its receptor involving RRLE in AD [23].
Conclusion
The findings of this study reveal that RRLE attenuates learning and memory deficits in 3xTg-AD mice, and thus, the plant material can potentially be developed as an alternative therapeutic agent for AD.
References
Archives


News Updates